Cephalothorax
- Crustaceans
Monolistra spp.
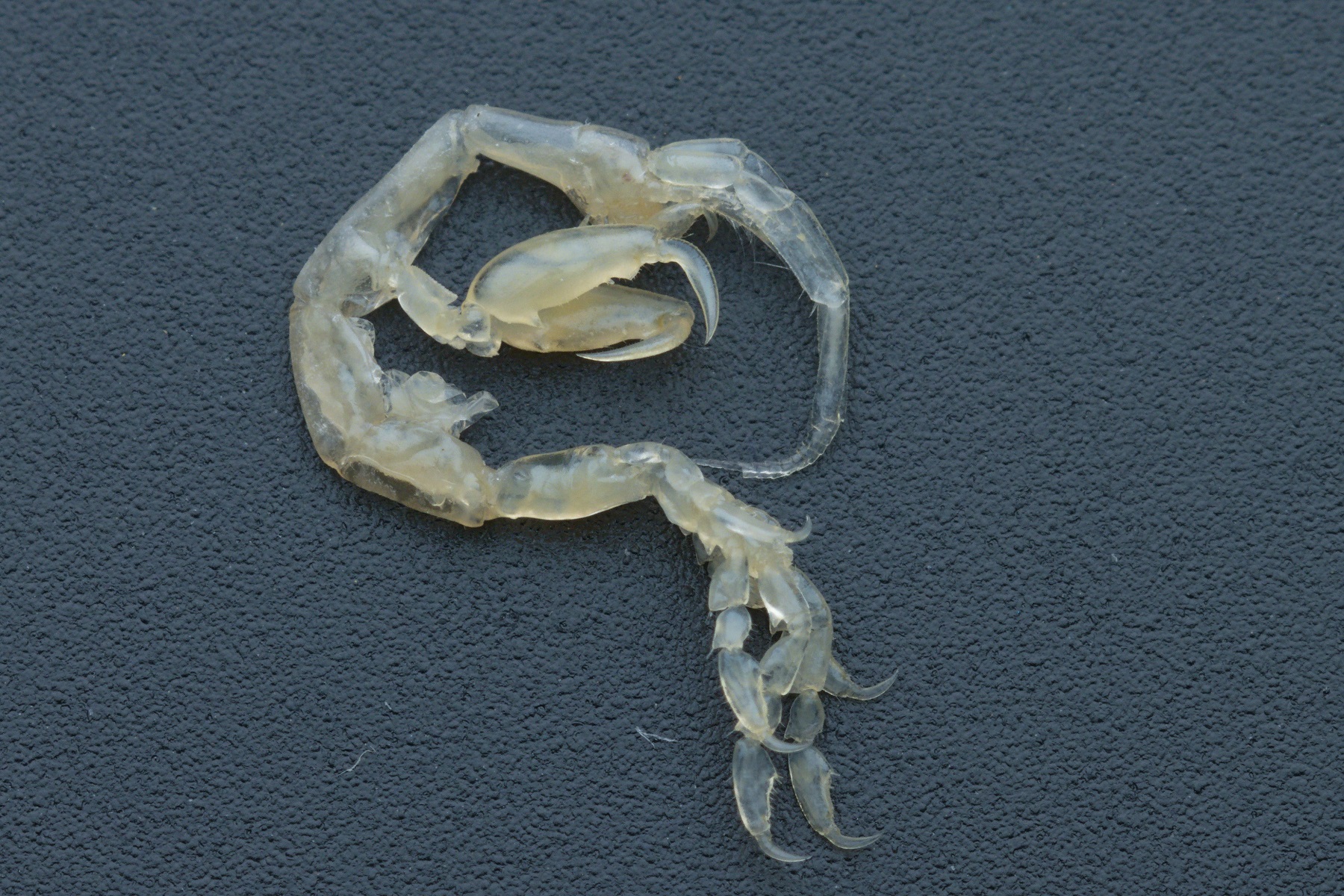
Monolistra isopods belong to the family of seapills (Sphaeromatidae). They are from one half to two centimeters large, have an arched body and are white or yellow due to the lack of pigment. They are capable of conglobation (rolling into a ball) and usually have the dorsal side of the body ornamented with intriguing protrusions in the form of bumps or curved spines of different lengths. The different species differ also in body size and uropod shape. Their body consists of a cephalothorax, pereon and pleon (abdomen). The cephalothorax and pleon are relatively small, with pereon being the most prominent body region. Pleon is usually highly arched.
The cephalothorax includes an acron, five head segments, and one additional segment. All the segments are fused into a uniform unit. The eyes are completely reduced. The head bears two pairs of antennae. Antennae II are a bit longer than the antennae I but never exceed one half of the body length. The antennae have a chemo- and mechanosensory functions. The mouthparts comprise a pair of mandibles, a pair of maxillae I, a pair of maxillae II, and a pair of maxillipeds. Evolutionary, the maxillipeds are modified appendages of the first thoracic segment.
The pereon consists of seven clearly delimited segments (pereomeres), each carrying a pair of appendages (pereopods). All the pereopods are similar, cylindrical walking legs. Only males of some species have subchelate second pair of pereopods. In females, a marsupium is developed at the ventral side of the pereon after the marsupial molt. The female lays fertilized eggs into the marsupium, which provides a safe environment for the development of offspring. The marsupium is basket-shaped and is formed by oostegites, which are leaflike outgrowths at the coxae of the second to fourth pair of pereopods.
The pleon is comprised of six body segments (pleomeres) and a telson. Due to the pleomere fusion the segmentation of pleon is obscured and appears tripartite. The first pleomere is free but hidden beneath the tergite of the seventh pereomere. The second and third pleomere are fused into a uniform structure. The last three pleomeres are fused with telson into a uniform pleotelson. The pleon carries five pairs of biramous, leaflike appendages (pleopods), which serve as gills and ventilate marsupium. In males the second pair of pleopods is modified as gonopods for the transfer of sperm during mating. The last, sixth pair of appendages on the pleon are uropods, which are differently shaped in different species. They can be completely reduced, short and stout, or long and slender.
The genus Monolistra comprises at least 47 species of troglobitic freshwater isopods, 33 of which already have a formal scientific description. In Slovenia 15 species are present, with one still lacking formal description. They are exclusively troglobitic, inhabiting underground waters of Dinaric Karst and south Alpine Karst. Some species are endemic, limited to just a few caves or springs and thus endangered. Presumably, the genus Monolistra dates back to Miocene when the European Continent and the Dinaric Alps were formed. They originate from marine ancestors and some of their relatives, such as Sphaeroma serratum, Lekanesphaera hookeri, and Dynamene bicolor, are present also in Slovenia. Marine ancestors of the genus Monolistra likely first inhabited surface freshwaters and only subsequently colonized groundwater. Another troglobitic genus belonging to Sphaeromatidae that inhabits European caves is Caecosphaeroma which comprises two species from France.
The isopods of genus Monolistra are detritivores feeding on decaying organic material, fungi, and bacteria. They move slowly and do not swim as do their marine relatives.
They exhibit three interesting anti-predator defense mechanisms. The first and the most obvious are the dorsal spines. Species with long spines usually co-occur with the largest groundwater predator, the olm (Proteus anguinus), while species without spines rarely do. The dorsal spines of Monolistra evolved more than once independently via convergent evolution, due to similar environmental factors. The second defense mechanism is conglobation, which allows the animal to hide its soft ventral side and be quickly washed away from the predator by the water current. The third defense mechanism is aggregation into groups by conspecifics, where an individual is less likely to become prey.
The reproduction and lifecycle of Monolistra isopods is relatively unknown. Species without dorsal spines, are known to exhibit amplexus (precopula) prior to mating, in which the male grasps the female from the dorsal side and guards it. In spiny species such amplexus is impossible and their mating behavior remains a mystery. In species without dorsal spines the males are generally larger than females, while the situation is reversed in species with dorsal spines. After successful mating the female lays bluish-green eggs into the marsupium. The number of eggs per clutch varies between species and differently sized females. The development of juveniles probably takes longer than in the marine relatives.
Different Monolistra species can co-occur in the same habitat and up to three species have been observed at the same locality. The co-occurring species differ in size and spine morphology, while the morphologically similar species are not found together. This indicates that the co-occurring species most likely occupy different ecological niches.
Chapter author: Žiga Fišer, Cene Fišer, Urban Bogataj
Photography: Teo Delić
3D model author: Urban Bogataj
More photos




Pereon
Pleon (abdomen)








The 3D Atlas of Arthropods project is funded by the University of Ljubljana and the Republic of Slovenia, the Ministry of Education, Science and Sport, and the European Union from the European Regional Development Fund.
Authors
- Urban Bogataj,
- Gregor Bračko,
- Teo Delič,
- Cene Fišer,
- Žiga Fišer,
- Rok Kostanjšek,
- Rudi Verovnik,
- Miloš Vittori,
- Valerija Zakšek.
Students Vito Ham, Vesna Jurjevič, Gaj Kušar, and Adrijan Samuel Stell Pičman also participated in the project.